MICROBIAL INFECTIONS OF HUMANS(HUMAN MICROBIOLOGY CONTD..)
VIRUS AS PARASITES:
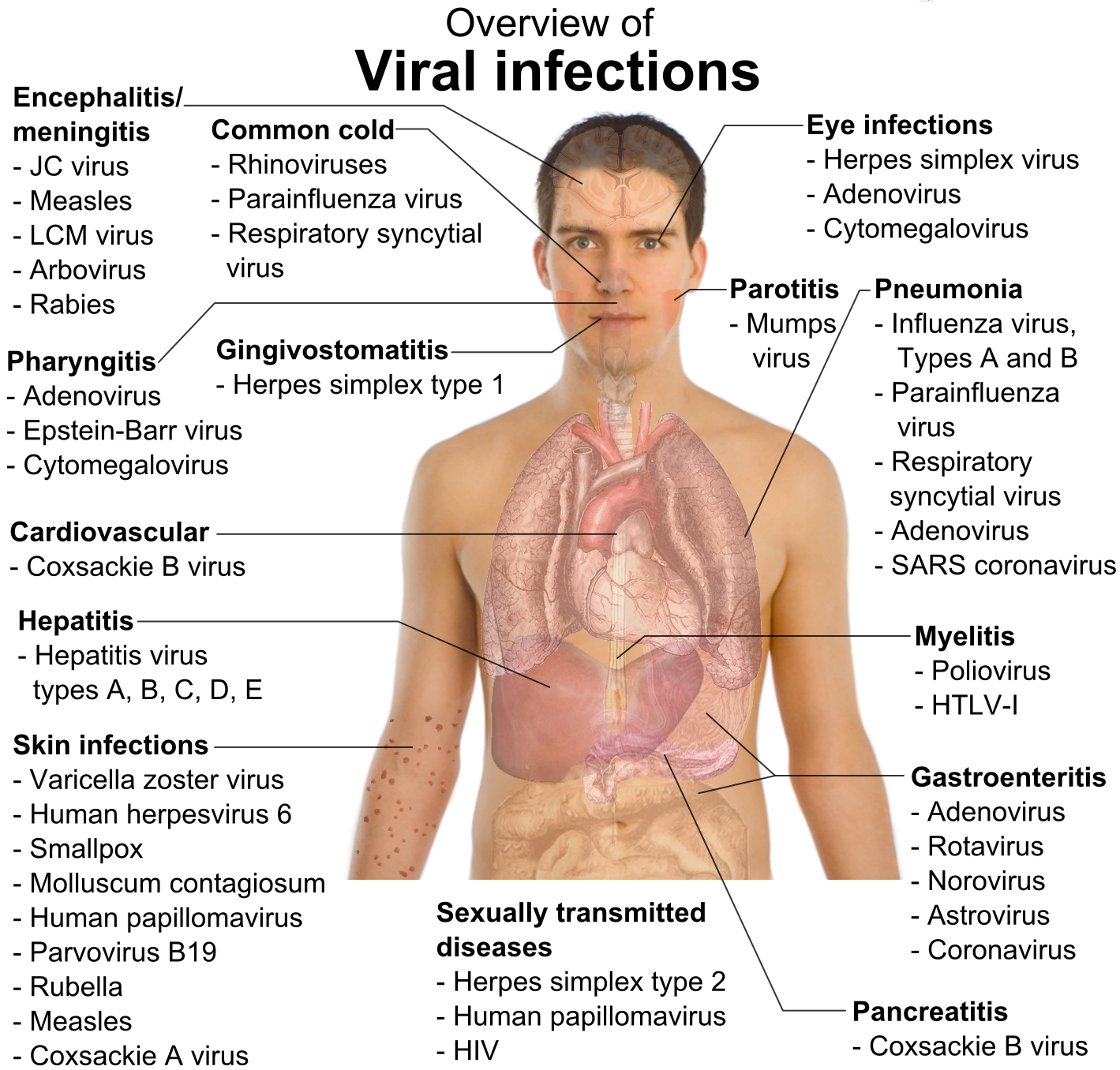
At a conservative estimate, there are at least thirty different viruses from fifteen taxonomic families that
are common causes of human illness. This number does not take into consideration the serological types
that exist in many of these viruses. So the common cold we are taking as one virus, whereas there are
over eighty serological types of rhinovirus. As every human will be infected by viruses in their lifetime,
we must concede that viruses are efficient parasites of humans. I am trying to outline some of the features of viruses that contribute to their success as human pathogens.
TYPES OF CLINICAL INFECTIONS:
The four broad types of viral infections recognised in humans. Because it is only those people with symptoms who come to our attention, it is easy to overlook the fact that most viral infections are asymptomatic. For the people who demonstrate clinical illness, the severity will range between mild and severe. The proportion of susceptible people who develop illness from those who are infected is called the attack rate, although it is not a true ‘rate’ and gives no indication of the total number of people infected.
The route of transmission does not fully account for the variation in attack rate between viruses, although clearly direct inoculation by parenteral transmission is likely to be more efficient in establishing an infection than transmission via aerosols. Unlike bacterial infections, many viral infections have a more damaging effect in older people compared to children. The childhood viral infections such as chickenpox and mumps are famously unforgiving in adults, whereas in children they cause rashes but otherwise usually fail to bother the child unduly. Why this should be is largely unknown.

Duration Extent Examples
Acute Local Rhinovirus,
Systemic Measles
Persistent Chronic Hepatitis B
Epstein Barr Virus
Latent Herpes Simplex Virus, Varicella zoster
Slow Bovine spongiform
encephalopathy
STRATEGIES FOR SUCCESSFUL VIRAL INFECTIONS:
Successful? By this we mean reproductive success: able to replicate and persist within a population. As
obligate intracellular parasites, viruses have an absolute requirement for their host. If the virus causes
damage or even death of the host it must do so after it has had sufficient time to manufacture high
numbers of new viruses and release them into the environment. Note also that whilst many bacteria can
multiply in the environment without the requirement of animal tissue, viruses can only multiply in a
host. Periods spent in the environment are simply a holding event for viruses in which no multiplication
can occur. Viral replication can be seen from a perspective in which the availability of the host
determines the strategy adopted for infecting humans. In dense populations rapid and short periods of
replication serve to transmit the virus such that new (and readily available) hosts becoming infected. In
sparsely populated areas, the virus will need to produce virus over a longer period so that the few new
hosts that come into contact with the infected host can be infected. This distinction is reflected in two
broad groupings of viral infections of humans: acute and persistent.
The acute infections rely on rapid multiplication of virus in order to manufacture and shed (transmit)
new virus before the host has had time to mount a neutralising immunity. Examples will be measles and
influenza viruses, both RNA viruses and ones that require sufficient numbers of susceptible hosts in
order to maintain themselves in the community. With the measles virus, humans appear to be the only
host. With influenza virus, however, a wide range of animals can be infected and the virus is therefore a
zoonosis, periodically sweeping into town as an epidemic having transferred from an animal reservoir.
Persistent infections in the host (not the environment) are those in which the virus is not eliminated but,
having established itself, will persist for the lifetime of the host. The virus is either replicating at a very
low level continually, such as chronic hepatitis virus infections, or is latent for most of the time but
periodically undergoes a short burst of full replication in which infectious virus is manufactured and
released, e.g. Herpes simplex in cold sores. Many persistent virus infections are acquired as children with little or no pathology and then persist for life. This lifetime of infection will optimise the
chances of transmission, preferably to other children. Persistent infections, by definition, require that the
virus is maintained within the host cell and thus seeks to avoid killing the cell (is not cytolytic).
Integration of the viral nucleic acid into the host chromosome is an appropriate means to help promote
long-term survival in the host and avoid contact with the immune system which will also seek to destroy
the infected cells. Whilst persistent virus infections are mostly DNA viruses such as Herpes and
papillomaviruses, retroviruses will insert themselves as proviral DNA into the host genome following
reverse transcription from RNA and many retrovirus infections are persistent.
The differences in acute and persistent infections have consequences for antiviral treatments. Specific
antiviral chemotherapy will only have significant effects on persistent infections where there is
continued production of virus. In acute infections, the virus is produced in greatest numbers just prior to
the development of symptoms. By the time the patient has sought help, the virus is no longer in full
manufacture.
Other differences result from the properties of DNA and RNA viruses. RNA viruses have a reduced gene capacity compared with DNA viruses. DNA viruses often possess sufficient genetic space (several hundred
genes) such that roughly 50 per cent code for proteins that can interfere with the host immunity. RNA viruses do not have sufficient gene space and utilise genetic variation in order to generate new variants of virus.
STAGES IN HUMAN INFECTIONS
The key stages in the process of an infection are:
• entry into host,
• primary replication,
• spread within host,
• exit from host,
• evade host response.
ENTRY INTO HOST
The routes of transmission will depend on the site of the body that represent the exit site for the infectious virus. The exit of virus from the body can occur via several vehicles: urine, faeces, semen,
saliva, blood, milk, skin, squamous epithelia. Hepatitis B virus is released into the bloodstream from the liver and the transfer of infected blood is the major vehicle for transmission. Examples include the
sharing of needles amongst drug abusers and the accidental transmission following blood transfusion.
With hepatitis B the transmission is parenteral and therefore the virus cannot be transmitted by sneezing
or touching. Note how the site of multiplication (the liver) need not be the infectious site (blood).
Conversely, rhinoviruses, the aetiological agents of the common cold, multiply in the upper respiratory
tract epithelia and are transmitted via the secretions from the runny nose.
There are several ways of listing routes of transmission of infectious agents . The traditional list is as follows:
• aerosol,
• faeco-oral,
• parenteral (including vertical transmission),
• sexually,
• direct contact.
The routes of horizontal transmission can be related to the sites in the human body that represent the
major sites of entry:
• skin,
• gastrointestinal tract,
• respiratory tract,
• genito-urinary tract.
This list can be sensibly reduced by grouping the last three into one to yield three key routes of entry for
a virus into a new human host:
• Entry through mucous membranes: respiratory, genitourinary and gastrointestinal tracts and the
conjunctiva.
• Entry through skin: insect bites (e.g. yellow fever virus); animal bites (e.g. rabies virus); skin to
skin (e.g. papillomavirus (warts, verrucas), Orf virus).
• Parenteral transmission: vertical transmission, direct inoculation into blood (needle, insect or
animal bite) or blood transfer through trauma (including sexual transmission).
Vertical transmission has clear advantages in that the foetus has the same blood supply as the mother
and has no defences against virus attack other than those present in the mother. One of the drawbacks of
vertical transmission is the risk of damaging the developing child in utero, i.e. foetal death and abortion.
A less extreme outcome is teratogenesis. Vertical transmission also includes transmission
via the germ line (i.e. through infected sperm or ovaries).
The properties of the virus will also influence those routes that are likely to be successful for the virus.
Viruses that cause gastroenteritis will need to resist exposure to gastric acid, bile acids and intestinal proteases if they are to reach the small intestine in any number and condition. As exemplified by rotaviruses, viruses causing gastroenteritis are not enveloped (which would be dissolved by bile acids) and tolerate acid pH and digestion by proteases. In fact, certain viruses are ‘activated’ by host proteases which act on surface proteins on the viral capsid. Numerous viruses that infect the respiratory tract are enveloped (notably influenza), reflecting the absence of lipid-dissolving secretions such as bile in the respiratory tract lining. For this reason the term ‘gastric flu’ is a misnomer. Influenza viruses cannot infect the stomach.
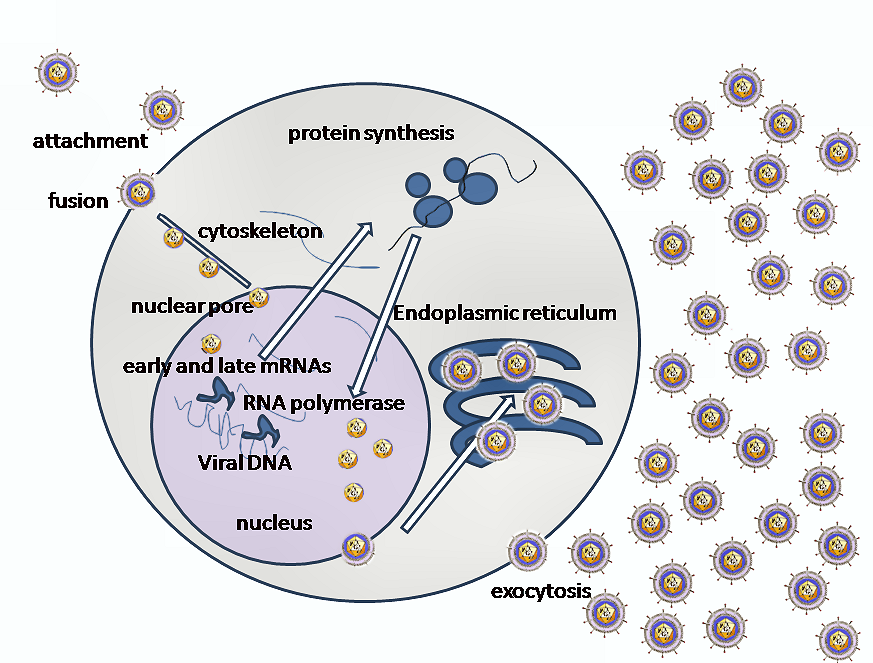
PRIMARY REPLICATION:
This depends on the site of entry: local infections will usually replicate at the site of entry. For example,
the rhinoviruses that cause the common cold will replicate in the epithelia in the nose and upper
respiratory tract. Local infections may invade the local lymph nodes but otherwise are restricted to the
site of entry. Systemic infections tend to have two-stage infection. The primary replication site serves as
an amplification of the virus from the small initial infectious dose and will be asymptomatic. Once the
virus has replicated at the local lymph node or epithelium, large numbers of virus are then released into
the bloodstream where they can attack the secondary tissues with corresponding tissue damage.
SPREAD WITHIN HOST:
The common cold will mostly restrict itself to the upper respiratory tract. Not all virus infections are
limited to localised infections but, instead, spread throughout the body (systemic infection) and have
secondary rounds of replication in other organs, yet not every cell in the body will succumb to viral
infection. The pathology and symptoms help distinguish different infections (e.g. rubella virus often
causes joint inflammation, whereas hepatitis viruses target the liver). The reason that viruses show such
tissue tropism(a restricted range of tissues that become infected) depends in part on the presence or
absence of appropriate receptor molecules on the host cell membranes and the relative permissiveness of
the cell type within that organ. This combination of factors means that the distribution of viral receptors
may be wider than any of the observed tissue tropism of the virus. Conversely, cell types other than
those in tissues displaying symptoms may be permissive for the virus but asymptomatic. Unless one
investigates all these other cells that may be infected, one can never be sure.
Many of the highly infectious virus infections of childhood (measles, chickenpox, rubella, mumps)
cause systemic infections reflecting rounds of viral replication in more than one organ of the body. The
virus enters at the appropriate site (skin, mucous membrane) and multiplies at the nearest available
tissue (e.g. epithelia). The virus spreads to the local lymph nodes from which it is released into the
bloodstream. This first viraemic phase carries the virus throughout the body and seeds the virus in
organs such as the liver, spleen and bone marrow. After a further round of multiplication, a second viraemic phase will deposit the virus in organs throughout the body. The second viraemia may be the
release of large numbers of virus into the bloodstream or the movement of virus within the cells of the
immune system (EBV, measles, HIV). The actual organ that is targeted from the secondary viraemia is
called the ‘target organ’ and will probably manifest itself in the symptoms. Only at this stage, however,
does overt disease become apparent with the patient showing signs and symptoms typical of the
infection in question. The childhood rashes seen with measles and chickenpox will appear following the
second viraemia. The exact cause of the viral rashes varies. Some rashes represent virus-infected cells being destroyed by the immune response, whereas others represent
blood vessel damage due to virus.
Having presented a generalised pattern of systemic viral infections, it is important to point out that
certain virus infections do not employ viraemic phases to spread through the body. Rabies virus moves
along the nerve fibres from the site of the bite towards the central nervous system. A significant
advantage of this pathway is the relative inaccessibility of the nervous system to immune surveillance
and the specific protection from the circulating antibodies, which cannot penetrate cells.
The different pathways taken by viruses in the course of an infection will often be reflected in the
incubation period. Local infections have shorter incubation periods (up to a week) whereas the systemic
infections incubate for 10 days or more. The important feature of this process is the long incubation
period during which the patient becomes infectious but shows no clinical disease. (The time from
between becoming infected and becoming infectious is called the ‘latent period’.) This is typical of
many viral infections, where the development of disease occurs after maximum numbers of virus are
being excreted. In this way transmission is favoured because the susceptible hosts are not running the
other way having seen the rash!
EXIT FROM HOST
What is the exit point from one host is likely to be the entry point for infection in the the next host. In
other words, the sites of viral replication will determine how and where the virus is transmitted to the
next host. Local infections will be shed from the local mucosa in secretions. The common cold is a
testament to the efficiency of the ‘secretions’. Blood-borne infections are that much more difficult to
transmit horizontally but nevertheless are able to escape through blood via insects, blood transfusion,
and trauma during sexual contact. A list of secretions that can contain viruses (saliva and nasal secretions, semen, milk, urine, vesicle fluids and faeces) does not take into account the duration of excretion of the virus, nor the numbers of virus, both of which can vary between virus depending on the strategy adopted. Is it better to shed large numbers of virus for a short while or smaller numbers over a longer period?
Cited By Kamal Singh Khadka
Msc Microbiology, TU.
Assistant Professor In Pokhara Bigyan Tatha Prabhidi Campus(PBPC), PU, PNC, LA, NA.
Pokhara, Nepal.
SOME SUGGESTED REFERENCES:
www.nativeremedies.com/ailment/types-of-viral-infections.html
www.merckmanuals.com/.../infections/viral_infections/overview_of_vira...
www.betterhealth.vic.gov.au/Bhcv2/.../Infections_bacterial_and_viral?...
kidshealth.org/parent/infections/
www.meningitis.org/disease-info/types-causes/viral-meningitis
www.nlm.nih.gov/medlineplus/viralinfections.html
www.betterhealth.vic.gov.au/Bhcv2/.../Infections_bacterial_and_viral?...
cmr.asm.org/content/23/1/74.full
www.ncbi.nlm.nih.gov/pubmed/12753686
www.patient.co.uk › Professional Reference
emedicine.medscape.com/article/211316-overview
en.wikipedia.org/wiki/HIV/AIDS
cid.oxfordjournals.org/content/32/6/897.full?linkType=FULL...







At a conservative estimate, there are at least thirty different viruses from fifteen taxonomic families that
are common causes of human illness. This number does not take into consideration the serological types
that exist in many of these viruses. So the common cold we are taking as one virus, whereas there are
over eighty serological types of rhinovirus. As every human will be infected by viruses in their lifetime,
we must concede that viruses are efficient parasites of humans. I am trying to outline some of the features of viruses that contribute to their success as human pathogens.
TYPES OF CLINICAL INFECTIONS:
The four broad types of viral infections recognised in humans. Because it is only those people with symptoms who come to our attention, it is easy to overlook the fact that most viral infections are asymptomatic. For the people who demonstrate clinical illness, the severity will range between mild and severe. The proportion of susceptible people who develop illness from those who are infected is called the attack rate, although it is not a true ‘rate’ and gives no indication of the total number of people infected.
The route of transmission does not fully account for the variation in attack rate between viruses, although clearly direct inoculation by parenteral transmission is likely to be more efficient in establishing an infection than transmission via aerosols. Unlike bacterial infections, many viral infections have a more damaging effect in older people compared to children. The childhood viral infections such as chickenpox and mumps are famously unforgiving in adults, whereas in children they cause rashes but otherwise usually fail to bother the child unduly. Why this should be is largely unknown.
Duration Extent Examples
Acute Local Rhinovirus,
Systemic Measles
Persistent Chronic Hepatitis B
Epstein Barr Virus
Latent Herpes Simplex Virus, Varicella zoster
Slow Bovine spongiform
encephalopathy
STRATEGIES FOR SUCCESSFUL VIRAL INFECTIONS:
Successful? By this we mean reproductive success: able to replicate and persist within a population. As
obligate intracellular parasites, viruses have an absolute requirement for their host. If the virus causes
damage or even death of the host it must do so after it has had sufficient time to manufacture high
numbers of new viruses and release them into the environment. Note also that whilst many bacteria can
multiply in the environment without the requirement of animal tissue, viruses can only multiply in a
host. Periods spent in the environment are simply a holding event for viruses in which no multiplication
can occur. Viral replication can be seen from a perspective in which the availability of the host
determines the strategy adopted for infecting humans. In dense populations rapid and short periods of
replication serve to transmit the virus such that new (and readily available) hosts becoming infected. In
sparsely populated areas, the virus will need to produce virus over a longer period so that the few new
hosts that come into contact with the infected host can be infected. This distinction is reflected in two
broad groupings of viral infections of humans: acute and persistent.
The acute infections rely on rapid multiplication of virus in order to manufacture and shed (transmit)
new virus before the host has had time to mount a neutralising immunity. Examples will be measles and
influenza viruses, both RNA viruses and ones that require sufficient numbers of susceptible hosts in
order to maintain themselves in the community. With the measles virus, humans appear to be the only
host. With influenza virus, however, a wide range of animals can be infected and the virus is therefore a
zoonosis, periodically sweeping into town as an epidemic having transferred from an animal reservoir.
Persistent infections in the host (not the environment) are those in which the virus is not eliminated but,
having established itself, will persist for the lifetime of the host. The virus is either replicating at a very
low level continually, such as chronic hepatitis virus infections, or is latent for most of the time but
periodically undergoes a short burst of full replication in which infectious virus is manufactured and
released, e.g. Herpes simplex in cold sores. Many persistent virus infections are acquired as children with little or no pathology and then persist for life. This lifetime of infection will optimise the
chances of transmission, preferably to other children. Persistent infections, by definition, require that the
virus is maintained within the host cell and thus seeks to avoid killing the cell (is not cytolytic).
Integration of the viral nucleic acid into the host chromosome is an appropriate means to help promote
long-term survival in the host and avoid contact with the immune system which will also seek to destroy
the infected cells. Whilst persistent virus infections are mostly DNA viruses such as Herpes and
papillomaviruses, retroviruses will insert themselves as proviral DNA into the host genome following
reverse transcription from RNA and many retrovirus infections are persistent.
The differences in acute and persistent infections have consequences for antiviral treatments. Specific
antiviral chemotherapy will only have significant effects on persistent infections where there is
continued production of virus. In acute infections, the virus is produced in greatest numbers just prior to
the development of symptoms. By the time the patient has sought help, the virus is no longer in full
manufacture.
Other differences result from the properties of DNA and RNA viruses. RNA viruses have a reduced gene capacity compared with DNA viruses. DNA viruses often possess sufficient genetic space (several hundred
genes) such that roughly 50 per cent code for proteins that can interfere with the host immunity. RNA viruses do not have sufficient gene space and utilise genetic variation in order to generate new variants of virus.
STAGES IN HUMAN INFECTIONS
The key stages in the process of an infection are:
• entry into host,
• primary replication,
• spread within host,
• exit from host,
• evade host response.
ENTRY INTO HOST
The routes of transmission will depend on the site of the body that represent the exit site for the infectious virus. The exit of virus from the body can occur via several vehicles: urine, faeces, semen,
saliva, blood, milk, skin, squamous epithelia. Hepatitis B virus is released into the bloodstream from the liver and the transfer of infected blood is the major vehicle for transmission. Examples include the
sharing of needles amongst drug abusers and the accidental transmission following blood transfusion.
With hepatitis B the transmission is parenteral and therefore the virus cannot be transmitted by sneezing
or touching. Note how the site of multiplication (the liver) need not be the infectious site (blood).
Conversely, rhinoviruses, the aetiological agents of the common cold, multiply in the upper respiratory
tract epithelia and are transmitted via the secretions from the runny nose.
There are several ways of listing routes of transmission of infectious agents . The traditional list is as follows:
• aerosol,
• faeco-oral,
• parenteral (including vertical transmission),
• sexually,
• direct contact.
The routes of horizontal transmission can be related to the sites in the human body that represent the
major sites of entry:
• skin,
• gastrointestinal tract,
• respiratory tract,
• genito-urinary tract.
This list can be sensibly reduced by grouping the last three into one to yield three key routes of entry for
a virus into a new human host:
• Entry through mucous membranes: respiratory, genitourinary and gastrointestinal tracts and the
conjunctiva.
• Entry through skin: insect bites (e.g. yellow fever virus); animal bites (e.g. rabies virus); skin to
skin (e.g. papillomavirus (warts, verrucas), Orf virus).
• Parenteral transmission: vertical transmission, direct inoculation into blood (needle, insect or
animal bite) or blood transfer through trauma (including sexual transmission).
Vertical transmission has clear advantages in that the foetus has the same blood supply as the mother
and has no defences against virus attack other than those present in the mother. One of the drawbacks of
vertical transmission is the risk of damaging the developing child in utero, i.e. foetal death and abortion.
A less extreme outcome is teratogenesis. Vertical transmission also includes transmission
via the germ line (i.e. through infected sperm or ovaries).
The properties of the virus will also influence those routes that are likely to be successful for the virus.
Viruses that cause gastroenteritis will need to resist exposure to gastric acid, bile acids and intestinal proteases if they are to reach the small intestine in any number and condition. As exemplified by rotaviruses, viruses causing gastroenteritis are not enveloped (which would be dissolved by bile acids) and tolerate acid pH and digestion by proteases. In fact, certain viruses are ‘activated’ by host proteases which act on surface proteins on the viral capsid. Numerous viruses that infect the respiratory tract are enveloped (notably influenza), reflecting the absence of lipid-dissolving secretions such as bile in the respiratory tract lining. For this reason the term ‘gastric flu’ is a misnomer. Influenza viruses cannot infect the stomach.
PRIMARY REPLICATION:
This depends on the site of entry: local infections will usually replicate at the site of entry. For example,
the rhinoviruses that cause the common cold will replicate in the epithelia in the nose and upper
respiratory tract. Local infections may invade the local lymph nodes but otherwise are restricted to the
site of entry. Systemic infections tend to have two-stage infection. The primary replication site serves as
an amplification of the virus from the small initial infectious dose and will be asymptomatic. Once the
virus has replicated at the local lymph node or epithelium, large numbers of virus are then released into
the bloodstream where they can attack the secondary tissues with corresponding tissue damage.
SPREAD WITHIN HOST:
The common cold will mostly restrict itself to the upper respiratory tract. Not all virus infections are
limited to localised infections but, instead, spread throughout the body (systemic infection) and have
secondary rounds of replication in other organs, yet not every cell in the body will succumb to viral
infection. The pathology and symptoms help distinguish different infections (e.g. rubella virus often
causes joint inflammation, whereas hepatitis viruses target the liver). The reason that viruses show such
tissue tropism(a restricted range of tissues that become infected) depends in part on the presence or
absence of appropriate receptor molecules on the host cell membranes and the relative permissiveness of
the cell type within that organ. This combination of factors means that the distribution of viral receptors
may be wider than any of the observed tissue tropism of the virus. Conversely, cell types other than
those in tissues displaying symptoms may be permissive for the virus but asymptomatic. Unless one
investigates all these other cells that may be infected, one can never be sure.
Many of the highly infectious virus infections of childhood (measles, chickenpox, rubella, mumps)
cause systemic infections reflecting rounds of viral replication in more than one organ of the body. The
virus enters at the appropriate site (skin, mucous membrane) and multiplies at the nearest available
tissue (e.g. epithelia). The virus spreads to the local lymph nodes from which it is released into the
bloodstream. This first viraemic phase carries the virus throughout the body and seeds the virus in
organs such as the liver, spleen and bone marrow. After a further round of multiplication, a second viraemic phase will deposit the virus in organs throughout the body. The second viraemia may be the
release of large numbers of virus into the bloodstream or the movement of virus within the cells of the
immune system (EBV, measles, HIV). The actual organ that is targeted from the secondary viraemia is
called the ‘target organ’ and will probably manifest itself in the symptoms. Only at this stage, however,
does overt disease become apparent with the patient showing signs and symptoms typical of the
infection in question. The childhood rashes seen with measles and chickenpox will appear following the
second viraemia. The exact cause of the viral rashes varies. Some rashes represent virus-infected cells being destroyed by the immune response, whereas others represent
blood vessel damage due to virus.
Having presented a generalised pattern of systemic viral infections, it is important to point out that
certain virus infections do not employ viraemic phases to spread through the body. Rabies virus moves
along the nerve fibres from the site of the bite towards the central nervous system. A significant
advantage of this pathway is the relative inaccessibility of the nervous system to immune surveillance
and the specific protection from the circulating antibodies, which cannot penetrate cells.
The different pathways taken by viruses in the course of an infection will often be reflected in the
incubation period. Local infections have shorter incubation periods (up to a week) whereas the systemic
infections incubate for 10 days or more. The important feature of this process is the long incubation
period during which the patient becomes infectious but shows no clinical disease. (The time from
between becoming infected and becoming infectious is called the ‘latent period’.) This is typical of
many viral infections, where the development of disease occurs after maximum numbers of virus are
being excreted. In this way transmission is favoured because the susceptible hosts are not running the
other way having seen the rash!
EXIT FROM HOST
What is the exit point from one host is likely to be the entry point for infection in the the next host. In
other words, the sites of viral replication will determine how and where the virus is transmitted to the
next host. Local infections will be shed from the local mucosa in secretions. The common cold is a
testament to the efficiency of the ‘secretions’. Blood-borne infections are that much more difficult to
transmit horizontally but nevertheless are able to escape through blood via insects, blood transfusion,
and trauma during sexual contact. A list of secretions that can contain viruses (saliva and nasal secretions, semen, milk, urine, vesicle fluids and faeces) does not take into account the duration of excretion of the virus, nor the numbers of virus, both of which can vary between virus depending on the strategy adopted. Is it better to shed large numbers of virus for a short while or smaller numbers over a longer period?
Cited By Kamal Singh Khadka
Msc Microbiology, TU.
Assistant Professor In Pokhara Bigyan Tatha Prabhidi Campus(PBPC), PU, PNC, LA, NA.
Pokhara, Nepal.
SOME SUGGESTED REFERENCES:
www.nativeremedies.com/ailment/types-of-viral-infections.html
www.merckmanuals.com/.../infections/viral_infections/overview_of_vira...
www.betterhealth.vic.gov.au/Bhcv2/.../Infections_bacterial_and_viral?...
kidshealth.org/parent/infections/
www.meningitis.org/disease-info/types-causes/viral-meningitis
www.nlm.nih.gov/medlineplus/viralinfections.html
www.betterhealth.vic.gov.au/Bhcv2/.../Infections_bacterial_and_viral?...
cmr.asm.org/content/23/1/74.full
www.ncbi.nlm.nih.gov/pubmed/12753686
www.patient.co.uk › Professional Reference
emedicine.medscape.com/article/211316-overview
en.wikipedia.org/wiki/HIV/AIDS
cid.oxfordjournals.org/content/32/6/897.full?linkType=FULL...

Comments