VIROLOGY
VIRUS IDENTIFICATION:
Identifying viral isolates is not an easy task. For one thing, viruses
cannot be seen at all without the use of an electron microscope.
Serological methods, such as Western blotting, are the most commonly used means of identification. In these tests, the virus is detected and identified by its reaction with antibodies.
Virologists can identify and characterize viruses by using
such modern molecular methods as use of restriction fragment
length polymorphisms (RFLPs) and the polymerase chain reaction(PCR).PCR PCR was used to amplify viral RNA to identify the west Nile virus in 1999 in the United States & SARS-associated coronavirus in china in 2002.
Viral Multiplication:
The nucleic acid in a virion contains only a few of the genes
needed for the synthesis of new viruses. These include genes for
the virion structural components, such as the capsid proteins,
and genes for a few of the enzymes used in the viral life cycle.
These enzymes are synthesized and functional only when the
virus is within the host cell. Viral enzymes are almost entirely
concerned with replicating or processing viral nucleic acid.
Enzymes needed for protein synthesis, ribosomes, tRNA, and energy production are supplied by the host cell and are used for
synthesizing viral proteins, including viral enzymes. Although
the smallest non enveloped virions do not contain any preformed
enzymes, the larger virions may contain one or a few enzymes,
which usually function in helping the virus penetrate the host
cell or replicate its own nucleic acid .
Thus, for a virus to multiply, it must invade a host cell and
take over the host's metabolic machinery. A single virion can give
rise to several or even thousands of similar viruses in a single
host cell. This process can drastically change the host cell and
usually causes its death. In a few viral infections, cells survive and
continue to produce viruses indefinitely.
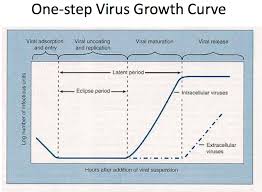
The multiplication of viruses can be demonstrated with
a one-step growth curve . The data are
obtained by infecting every cell in a culture and then testing the
culture medium and cells for virions and viral proteins and nucleic acids.
Multiplication of Bacteriophages:
Although the means by which a virus enters and exits a host cell
may vary, the basic mechanism of viral multiplication is similar
for all viruses. Bacteriophages can multiply by two alternative
mechanisms: the lytic cycle or the lysogenic cycle. The lytic cycle
ends with the lysis and death of the host cell, whereas the host
cell remains alive in the lysogenic cycle. Because the T-even bacteriophages
(T2, 14, and T6) have been studied most extensively,
we will describe the multiplication of T-even bacteriophages in
their host, E. coli, as an example of the lytic cycle.
T-Even Bacteriophages: The Lytic Cycle:
The virions of T-even bacteriophages are large, complex, and
non enveloped, with a char
 acteristic head -and -tail structure. The length of DNA contained
acteristic head -and -tail structure. The length of DNA contained
in these bacteriophages is only abou l 6% of that contained
in E. coli, yet the phage has enough DNA for over 100 genes. The
multiplication cycle of these phages, like that of all viruses,
occurs in five distinct stages: attachment, penetration, biosynthesis maturation, and release.

Step 1: Attachment: After a chance collision between phage particles
and bacteria, attachment , or adsorption, occurs. During this
process, an attachment site on the virus attaches to a complementary
receptor site on the bacterial cell. This attachment is a
chemical interaction in which weak bonds are formed between
the attachment and receptor sites. T-even bacteriophages use
fibers at the end of the tail as attachment sites. The complementary
receptor sites are on the bacterial cell wall.
Step 2: Penetration: After attachment, the T-even bacteriophage
injects its DNA (nucleic acid) into the bacterium. To do this, the
bacteriophage's tail releases an enzyme, phage lysozyme, which
breaks down a portion of the bacterial cell wall. During the
process of penetration, the tail sheath of the phage contracts, and
the tail core is driven through the cell wall. When the tip of the
core reaches the plasma membrane, the DNA from the bacteriophage's
head passes through the tail core, through the plasma
membrane, and enters the bacterial cell. The capsid remains outside
the bacterial cell. Therefore, the phage particle functions like
a hypodermic syringe to inject its DNA into the bacterial cell.
Step 3: Biosynthesis: Once the bacteriophage DNA has reached the
cytoplasm of the host cell, the biosynthesis of viral nucleic acid
and protein occurs. Host protein synthesis is stopped by virus induced
degradation of the host DNA, viral proteins that interfere
with transcription, or the repression of translation.
Initially, the phage uses the host cell's nucleotides and several
of its enzymes to synthesize many copies of phage DNA. Soon
after, the biosynthesis of viral proteins begins. Any RNA transcribed
in the cell is mRNA transcribed from phage DNA for the
biosynthesis of phage enzymes and capsid proteins. The host
cell's ribosomes, enzymes, and amino adds are used for translation.
Genetic controls regulate when different regions of phage
DNA are transcribed into mRNA during the multiplication
cycle. For example, early messages are translated into early phage
proteins, the enzymes used in the synthesis of phage DNA. Also,
late messages are translated into late phage proteins for the synthesis
of capsid proteins.
For several minutes following infection, complete phages
cannot be found in the host cell. Only separate components-
DNA and protein- can be detected. The period during viral
multiplication when complete, infective virions are not yet present is called eclipse period.
Step 4: Maturation:
In the next sequence of events, maturation occurs.
In this process, bacteriophage DNA and capsids are assembled
into complete virions. The viral components essentially assemble
into a viral particle spontaneously, eliminating the need for many
nonstructural genes and gene products. The phage heads and tails
are separately assembled from protein subunits, and the head is
filled with phage DNA and attached to the tail.
Step 5: Release or Lysis:
The final stage of viral multiplication is the release of
virions from the host cell. The term lysis is generally used for this
stage in the multiplication of T-even phages because in this case, the
plasma membrane actually breaks open (lyses). Lysozyme, which is
encoded by a phage gene, is synthesized within the cell. This
enzyme causes the bacterial cell wall to break down, and the newly
produced bacteriophages are released from the host cell. The
released bacteriophages infect other susceptible cells in the vicinity,
and the viral multiplication cycle is repeated within those cells.
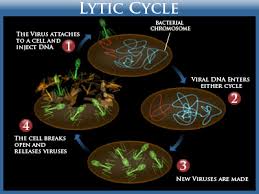
Fig: Lytic Cycle
Bacteriophage Lambda (A.): The Lysogenic Cycle:
In contrast to T-even bacteriophages, some viruses do not cause
lysis and death of the host cell when they multiply. These
lysogenic phages (also called temperate phages) may indeed proceed
through a lytic cycle, but they are also capable of incorporating
their DNA into the host cell's DNA to begin a lysogenic
cycle. In lysogeny, the phage remains latent (inactive). The participating
bacterial host cells are known as lysogenic cells.
We will use the bacteriophage A (lambda), a well-studied lysogenic
phage, as an example of the lysogenic cycle.
Upon penetration into an E. coli cell,
a the originally linear phage DNA forms a circle.
¢\ This circle can multiply and be transcribed,
Qi) leading to the production of new phage and to cell lysis (the
lytic cycle).
€Ii) Alternatively, the circle can recombine with and become
part of the circular bacterial DNA (the lysogenic cycle). The
inserted phage DNA is now called a prophage. Most of the
prophage genes are repressed by two repressor proteins that
are the products of phage genes. These repressors stop transcription
of all the other phage genes by binding to operators.
Thus, the phage genes that would otherwise direct the
synthesis and release of new virions are turned off, in much
the same way that the genes of the E. coli lac operon are
turned off by the lac repressor .
Every time the host cell's machinery replicates the bacterial
chromosome,
e it also replicates the prophage DNA. The prophage remains
latent within the progeny cells.
o However, a rare spontaneous event, or the action of UV light
or certain chemicals, can lead to the excision (popping-out)
of the phage DNA, and to initiation of the lytic cycle.
There are three important results of lysogeny. First, the lysogenic
cells are immune to reinfection by the same phage.
(However, the host cell is not immune to infection by other
phage types. ) The second result of lysogeny is phage conversion;
that is, the host cell may exhibit new properties. For example, the
bacterium Corynebacterium diphtheriae, which causes diphtheria,
is a pathogen whose disease-producing properties are related
to the synthesis of a toxin . The organism can produce toxin only
when it carries a lysogenic phage, because the prophage carries
the gene coding for the toxin . As another example, only streptococci
carrying a lysogenic phage are capable of causing toxic
shock syndrome. The toxin produced by Clostridium botulinum,
which causes botulism, is encoded by a prophage gene, as is the
Shiga toxin produced by pathogenic strains of E. coli.
The third result of lysogeny is that it makes specialized
transduction possible. Recall from Chapter 8 that bacterial
genes can be picked up in a phage coat and transferred to another
bacterium in a process called generalized transduction . Any bacterial genes can be transferred by
generalized transduction because the host chromosome is broken
down into fragments, any of which can be packaged into a
phage coat. In specialized transduction, however, only certain bacterial genes can be transferred.
Specialized transduction is mediated by a lysogenic phage,
which packages bacterial DNA along with its own DNA in the same capsid. When a prophage is excised from the host chromosome,
adjacent genes from either side may remain attached to the
phage DNA.The phage carries this gene to a galactose-negative cell,
which then becomes galactose-positive.
Certain animal viruses can undergo processes very similar to
lysogeny. Animal viruses that can remain latent in cells for long
periods without multiplying or causing disease may become
inserted into a host chromosome or remain separate from host
DNA in a repressed state (as some lysogenic phages).

Fig: Lysogenic Cycle Of Bacteriophage Lambda E.coli

Fig: Lytic vs Lysogenic Cycle
Cited By Kamal Singh Khadka
Msc Microbiology TU
&
Anil Bhujel
Bsc Microbiology, TU
Identifying viral isolates is not an easy task. For one thing, viruses
cannot be seen at all without the use of an electron microscope.
Serological methods, such as Western blotting, are the most commonly used means of identification. In these tests, the virus is detected and identified by its reaction with antibodies.
Virologists can identify and characterize viruses by using
such modern molecular methods as use of restriction fragment
length polymorphisms (RFLPs) and the polymerase chain reaction(PCR).PCR PCR was used to amplify viral RNA to identify the west Nile virus in 1999 in the United States & SARS-associated coronavirus in china in 2002.
Viral Multiplication:
The nucleic acid in a virion contains only a few of the genes
needed for the synthesis of new viruses. These include genes for
the virion structural components, such as the capsid proteins,
and genes for a few of the enzymes used in the viral life cycle.
These enzymes are synthesized and functional only when the
virus is within the host cell. Viral enzymes are almost entirely
concerned with replicating or processing viral nucleic acid.
Enzymes needed for protein synthesis, ribosomes, tRNA, and energy production are supplied by the host cell and are used for
synthesizing viral proteins, including viral enzymes. Although
the smallest non enveloped virions do not contain any preformed
enzymes, the larger virions may contain one or a few enzymes,
which usually function in helping the virus penetrate the host
cell or replicate its own nucleic acid .
Thus, for a virus to multiply, it must invade a host cell and
take over the host's metabolic machinery. A single virion can give
rise to several or even thousands of similar viruses in a single
host cell. This process can drastically change the host cell and
usually causes its death. In a few viral infections, cells survive and
continue to produce viruses indefinitely.
The multiplication of viruses can be demonstrated with
a one-step growth curve . The data are
obtained by infecting every cell in a culture and then testing the
culture medium and cells for virions and viral proteins and nucleic acids.
Multiplication of Bacteriophages:
Although the means by which a virus enters and exits a host cell
may vary, the basic mechanism of viral multiplication is similar
for all viruses. Bacteriophages can multiply by two alternative
mechanisms: the lytic cycle or the lysogenic cycle. The lytic cycle
ends with the lysis and death of the host cell, whereas the host
cell remains alive in the lysogenic cycle. Because the T-even bacteriophages
(T2, 14, and T6) have been studied most extensively,
we will describe the multiplication of T-even bacteriophages in
their host, E. coli, as an example of the lytic cycle.
T-Even Bacteriophages: The Lytic Cycle:
The virions of T-even bacteriophages are large, complex, and
non enveloped, with a char
in these bacteriophages is only abou l 6% of that contained
in E. coli, yet the phage has enough DNA for over 100 genes. The
multiplication cycle of these phages, like that of all viruses,
occurs in five distinct stages: attachment, penetration, biosynthesis maturation, and release.
Step 1: Attachment: After a chance collision between phage particles
and bacteria, attachment , or adsorption, occurs. During this
process, an attachment site on the virus attaches to a complementary
receptor site on the bacterial cell. This attachment is a
chemical interaction in which weak bonds are formed between
the attachment and receptor sites. T-even bacteriophages use
fibers at the end of the tail as attachment sites. The complementary
receptor sites are on the bacterial cell wall.
Step 2: Penetration: After attachment, the T-even bacteriophage
injects its DNA (nucleic acid) into the bacterium. To do this, the
bacteriophage's tail releases an enzyme, phage lysozyme, which
breaks down a portion of the bacterial cell wall. During the
process of penetration, the tail sheath of the phage contracts, and
the tail core is driven through the cell wall. When the tip of the
core reaches the plasma membrane, the DNA from the bacteriophage's
head passes through the tail core, through the plasma
membrane, and enters the bacterial cell. The capsid remains outside
the bacterial cell. Therefore, the phage particle functions like
a hypodermic syringe to inject its DNA into the bacterial cell.
Step 3: Biosynthesis: Once the bacteriophage DNA has reached the
cytoplasm of the host cell, the biosynthesis of viral nucleic acid
and protein occurs. Host protein synthesis is stopped by virus induced
degradation of the host DNA, viral proteins that interfere
with transcription, or the repression of translation.
Initially, the phage uses the host cell's nucleotides and several
of its enzymes to synthesize many copies of phage DNA. Soon
after, the biosynthesis of viral proteins begins. Any RNA transcribed
in the cell is mRNA transcribed from phage DNA for the
biosynthesis of phage enzymes and capsid proteins. The host
cell's ribosomes, enzymes, and amino adds are used for translation.
Genetic controls regulate when different regions of phage
DNA are transcribed into mRNA during the multiplication
cycle. For example, early messages are translated into early phage
proteins, the enzymes used in the synthesis of phage DNA. Also,
late messages are translated into late phage proteins for the synthesis
of capsid proteins.
For several minutes following infection, complete phages
cannot be found in the host cell. Only separate components-
DNA and protein- can be detected. The period during viral
multiplication when complete, infective virions are not yet present is called eclipse period.
Step 4: Maturation:
In the next sequence of events, maturation occurs.
In this process, bacteriophage DNA and capsids are assembled
into complete virions. The viral components essentially assemble
into a viral particle spontaneously, eliminating the need for many
nonstructural genes and gene products. The phage heads and tails
are separately assembled from protein subunits, and the head is
filled with phage DNA and attached to the tail.
Step 5: Release or Lysis:
The final stage of viral multiplication is the release of
virions from the host cell. The term lysis is generally used for this
stage in the multiplication of T-even phages because in this case, the
plasma membrane actually breaks open (lyses). Lysozyme, which is
encoded by a phage gene, is synthesized within the cell. This
enzyme causes the bacterial cell wall to break down, and the newly
produced bacteriophages are released from the host cell. The
released bacteriophages infect other susceptible cells in the vicinity,
and the viral multiplication cycle is repeated within those cells.
Fig: Lytic Cycle
Bacteriophage Lambda (A.): The Lysogenic Cycle:
In contrast to T-even bacteriophages, some viruses do not cause
lysis and death of the host cell when they multiply. These
lysogenic phages (also called temperate phages) may indeed proceed
through a lytic cycle, but they are also capable of incorporating
their DNA into the host cell's DNA to begin a lysogenic
cycle. In lysogeny, the phage remains latent (inactive). The participating
bacterial host cells are known as lysogenic cells.
We will use the bacteriophage A (lambda), a well-studied lysogenic
phage, as an example of the lysogenic cycle.
Upon penetration into an E. coli cell,
a the originally linear phage DNA forms a circle.
¢\ This circle can multiply and be transcribed,
Qi) leading to the production of new phage and to cell lysis (the
lytic cycle).
€Ii) Alternatively, the circle can recombine with and become
part of the circular bacterial DNA (the lysogenic cycle). The
inserted phage DNA is now called a prophage. Most of the
prophage genes are repressed by two repressor proteins that
are the products of phage genes. These repressors stop transcription
of all the other phage genes by binding to operators.
Thus, the phage genes that would otherwise direct the
synthesis and release of new virions are turned off, in much
the same way that the genes of the E. coli lac operon are
turned off by the lac repressor .
Every time the host cell's machinery replicates the bacterial
chromosome,
e it also replicates the prophage DNA. The prophage remains
latent within the progeny cells.
o However, a rare spontaneous event, or the action of UV light
or certain chemicals, can lead to the excision (popping-out)
of the phage DNA, and to initiation of the lytic cycle.
There are three important results of lysogeny. First, the lysogenic
cells are immune to reinfection by the same phage.
(However, the host cell is not immune to infection by other
phage types. ) The second result of lysogeny is phage conversion;
that is, the host cell may exhibit new properties. For example, the
bacterium Corynebacterium diphtheriae, which causes diphtheria,
is a pathogen whose disease-producing properties are related
to the synthesis of a toxin . The organism can produce toxin only
when it carries a lysogenic phage, because the prophage carries
the gene coding for the toxin . As another example, only streptococci
carrying a lysogenic phage are capable of causing toxic
shock syndrome. The toxin produced by Clostridium botulinum,
which causes botulism, is encoded by a prophage gene, as is the
Shiga toxin produced by pathogenic strains of E. coli.
The third result of lysogeny is that it makes specialized
transduction possible. Recall from Chapter 8 that bacterial
genes can be picked up in a phage coat and transferred to another
bacterium in a process called generalized transduction . Any bacterial genes can be transferred by
generalized transduction because the host chromosome is broken
down into fragments, any of which can be packaged into a
phage coat. In specialized transduction, however, only certain bacterial genes can be transferred.
Specialized transduction is mediated by a lysogenic phage,
which packages bacterial DNA along with its own DNA in the same capsid. When a prophage is excised from the host chromosome,
adjacent genes from either side may remain attached to the
phage DNA.The phage carries this gene to a galactose-negative cell,
which then becomes galactose-positive.
Certain animal viruses can undergo processes very similar to
lysogeny. Animal viruses that can remain latent in cells for long
periods without multiplying or causing disease may become
inserted into a host chromosome or remain separate from host
DNA in a repressed state (as some lysogenic phages).
Fig: Lysogenic Cycle Of Bacteriophage Lambda E.coli
Fig: Lytic vs Lysogenic Cycle
Cited By Kamal Singh Khadka
Msc Microbiology TU
&
Anil Bhujel
Bsc Microbiology, TU

Comments