MICROBIAL MOLECULAR BIOLOGY AND GENETICS
Microbial Recombination and Plasmids:
Mapping the Genome:
Finding the location of genes in any organism’s genome is a very
complex task. This section surveys approaches to mapping the
bacterial genome, using E.coli as an example. All three modes of
gene transfer and recombination have been used in mapping.
Mapping the Genome:
Finding the location of genes in any organism’s genome is a very
complex task. This section surveys approaches to mapping the
bacterial genome, using E.coli as an example. All three modes of
gene transfer and recombination have been used in mapping.
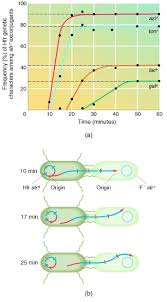
Hfr conjugation is frequently used to map the relative location of bacterial genes. This technique rests on the observation
that during conjugation the linear chromosome moves from
donor to recipient at a constant rate. In an interrupted mating
experiment the conjugation bridge is broken and Hfr F mating is stopped at various intervals after the start of conjugation by
mixing the culture vigorously in a blender. The
order and timing of gene transfer can be determined because they
are a direct reflection of the order of genes on the bacterial chromosome. For example, extrapolation of the curves in figure back to the x-axis will give the time at which
each gene just began to enter the recipient. The result is a circular chromosome map with distances expressed in terms of the
minutes elapsed until a gene is transferred. This technique can
fairly precisely locate genes 3 minutes or more apart. The heights
of the plateaus in figure bare lower for genes that are more
distant from the F factor (the origin of transfer) because there is
an ever-greater chance that the conjugation bridge will spontaneously break as transfer continues. Because of the relatively
large size of the E. coli genome, it is not possible to generate a
map from one Hfr strain. Therefore several Hfr strains with the F
plasmid integrated at different locations must be used and their
maps superimposed on one another. The overall map is adjusted
to 100 minutes, although complete transfer may require somewhat more than 100 minutes. In a sense, minutes are an indication
of map distance and not strictly a measure of time. Zero time is
set at the threonine (thr) locus.

 Gene linkage, or the proximity of two genes on a chromosome, also can be determined from transformation by measuring
Gene linkage, or the proximity of two genes on a chromosome, also can be determined from transformation by measuring
the frequency with which two or more genes simultaneously
transform a recipient cell. Consider the case for cotransformation
by two genes. In theory, a bacterium could simultaneously receive
two genes, each carried on a separate DNA fragment. However, it
is much more likely that the genes reside on the same fragment.
If two genes are closely linked on the chromosome, then they
should be able to cotransform. The closer the genes are together,
the more often they will be carried on the same fragment and the
higher will be the frequency of cotransformation. If genes are
spaced a great distance apart, they will be carried on separate
DNA fragments and the frequency of double transformants will
equal the product of the individual transformation frequencies.
Generalized transduction can be used to obtain linkage information in much the same way as transformation. Linkages usually
are expressed as cotransduction frequencies, using the argument
that the closer two genes are to each other, the more likely they
both will reside on the DNA fragment incorporated into a single
phage capsid. The E. coliphage P1 is often used in such mapping
because it can randomly transduce up to 1 to 2% of the genome.
Specialized transduction is used to find which phage attachment site is close to a specific gene. The relative locations of specific phage attsites are known from conjugational mapping, and
the genes linked to each attsite can be determined by means of
specialized transduction. These data allow precise placement of
genes on the chromosome.
A simplified genetic map of E. coli K12 is given in figure below.
Because conjugation data are not high resolution and cannot be used
to position genes that are very close together, the whole map is
developed using several mapping techniques. Usually, interrupted
mating data are combined with those from cotransduction and cotransformation studies. Data from recombination studies also are
used. Normally a new genetic marker in the E. coli genome is located within a relatively small region of the genome (10 to 15 minutes long) using a series of Hfr strains with F factor integration sites
scattered throughout the genome. Once the genetic marker has been
located with respect to several genes in the same region, its position
relative to nearby neighbors is more accurately determined using
transformation and transduction studies. Recent maps of the E. coli
chromosome give the locations of more than a thousand genes. Remember that the genetic map only depicts physical reality in a relative sense. A map unit in one region of the genome may not be the same physical distance as a unit in another part .
Genetic maps provide useful information in addition to the
order of the genes. For example, there is considerable clustering
of genes in E. coli K12 . In the regions around 2, 17,
and 27 minutes, there are many genes, whereas relatively few genetic markers are found in the 33 minute region. The areas apparently lacking genes may well have undiscovered genes, but
perhaps their function is not primarily that of coding genetic information. One hypothesis is that the 33 minute region is involved
in attachment of the E. coli chromosome to the plasma membrane
during replication and cell division. It is interesting that this region is almost exactly opposite the origin of replication for chromosome (Oric).
Cited By Kamal Singh Khadka
Msc Microbiology, TU
Assistant Professor In Pokhara University, Regional College Of Science & Technology, PNC, LA ,NA.
Pokhara, Nepal

Fig: E.coli K12 Genetic Map
Please Refer This Links To:
en.wikipedia.org/wiki/Human_Genome_Project
en.wikipedia.org/wiki/Gene_mapping
science.howstuffworks.com/life/genetic/designer-children1.htm
www.huffingtonpost.com/.../human-genome-project_b_3081121.html
www.genomenewsnetwork.org/resources/whats...genome/Chp3_1.shtml
www.genome.gov › Education › Fact Sheets
each gene just began to enter the recipient. The result is a circular chromosome map with distances expressed in terms of the
minutes elapsed until a gene is transferred. This technique can
fairly precisely locate genes 3 minutes or more apart. The heights
of the plateaus in figure bare lower for genes that are more
distant from the F factor (the origin of transfer) because there is
an ever-greater chance that the conjugation bridge will spontaneously break as transfer continues. Because of the relatively
large size of the E. coli genome, it is not possible to generate a
map from one Hfr strain. Therefore several Hfr strains with the F
plasmid integrated at different locations must be used and their
maps superimposed on one another. The overall map is adjusted
to 100 minutes, although complete transfer may require somewhat more than 100 minutes. In a sense, minutes are an indication
of map distance and not strictly a measure of time. Zero time is
set at the threonine (thr) locus.
the frequency with which two or more genes simultaneously
transform a recipient cell. Consider the case for cotransformation
by two genes. In theory, a bacterium could simultaneously receive
two genes, each carried on a separate DNA fragment. However, it
is much more likely that the genes reside on the same fragment.
If two genes are closely linked on the chromosome, then they
should be able to cotransform. The closer the genes are together,
the more often they will be carried on the same fragment and the
higher will be the frequency of cotransformation. If genes are
spaced a great distance apart, they will be carried on separate
DNA fragments and the frequency of double transformants will
equal the product of the individual transformation frequencies.
Generalized transduction can be used to obtain linkage information in much the same way as transformation. Linkages usually
are expressed as cotransduction frequencies, using the argument
that the closer two genes are to each other, the more likely they
both will reside on the DNA fragment incorporated into a single
phage capsid. The E. coliphage P1 is often used in such mapping
because it can randomly transduce up to 1 to 2% of the genome.
Specialized transduction is used to find which phage attachment site is close to a specific gene. The relative locations of specific phage attsites are known from conjugational mapping, and
the genes linked to each attsite can be determined by means of
specialized transduction. These data allow precise placement of
genes on the chromosome.
A simplified genetic map of E. coli K12 is given in figure below.
Because conjugation data are not high resolution and cannot be used
to position genes that are very close together, the whole map is
developed using several mapping techniques. Usually, interrupted
mating data are combined with those from cotransduction and cotransformation studies. Data from recombination studies also are
used. Normally a new genetic marker in the E. coli genome is located within a relatively small region of the genome (10 to 15 minutes long) using a series of Hfr strains with F factor integration sites
scattered throughout the genome. Once the genetic marker has been
located with respect to several genes in the same region, its position
relative to nearby neighbors is more accurately determined using
transformation and transduction studies. Recent maps of the E. coli
chromosome give the locations of more than a thousand genes. Remember that the genetic map only depicts physical reality in a relative sense. A map unit in one region of the genome may not be the same physical distance as a unit in another part .
Genetic maps provide useful information in addition to the
order of the genes. For example, there is considerable clustering
of genes in E. coli K12 . In the regions around 2, 17,
and 27 minutes, there are many genes, whereas relatively few genetic markers are found in the 33 minute region. The areas apparently lacking genes may well have undiscovered genes, but
perhaps their function is not primarily that of coding genetic information. One hypothesis is that the 33 minute region is involved
in attachment of the E. coli chromosome to the plasma membrane
during replication and cell division. It is interesting that this region is almost exactly opposite the origin of replication for chromosome (Oric).
Cited By Kamal Singh Khadka
Msc Microbiology, TU
Assistant Professor In Pokhara University, Regional College Of Science & Technology, PNC, LA ,NA.
Pokhara, Nepal

Fig: E.coli K12 Genetic Map
Please Refer This Links To:
en.wikipedia.org/wiki/Human_Genome_Project
en.wikipedia.org/wiki/Gene_mapping
science.howstuffworks.com/life/genetic/designer-children1.htm
www.huffingtonpost.com/.../human-genome-project_b_3081121.html
www.genomenewsnetwork.org/resources/whats...genome/Chp3_1.shtml
www.genome.gov › Education › Fact Sheets

Comments