MICROBIAL MOLECULAR BIOLOGY AND GENETICS
Microbial Recombination and Plasmids:
Transposable Elements:
The chromosomes of bacteria, viruses, and eucaryotic cells contain pieces of DNA that move around the genome. Such movement
is called transposition. DNA segments that carry the genes required for this process and consequently move about chromosomes are transposable elements or transposons.Unlike other
processes that reorganize DNA, transposition does not require extensive areas of homology between the transposon and its destination site. Transposons behave somewhat like lysogenic prophages
except that they originate in one chromosomal
location and can move to a different location in the same chromosome. Transposable elements differ from phages in lacking a virus
life cycle and from plasmids in being unable to reproduce autonomously and to exist apart from the chromosome. They were
first discovered in the 1940s by Barbara McClintock during her
studies on maize genetics (a discovery that won her the Nobel
prize in 1983). They have been most intensely studied in bacteria.
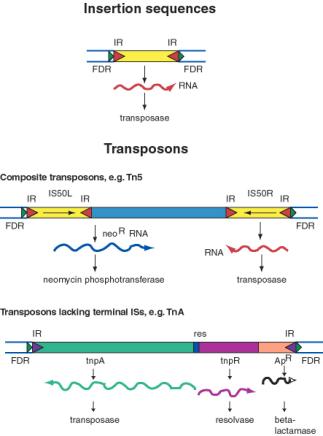
The simplest transposable elements are insertion sequences
or IS elements . An IS element is a short sequence
of DNA (around 750 to 1,600 base pairs [bp] in length) containing only the genes for those enzymes required for its transposition and bounded at both ends by identical or very similar sequences of nucleotides in reversed orientation known as inverted
repeats . Inverted repeats are usually about 15 to 25
base pairs long and vary among IS elements so that each type of
IS has its own characteristic inverted repeats. Between the inverted repeats is a gene that codes for an enzyme called transposase(and sometimes a gene for another essential protein). This
enzyme is required for transposition and accurately recognizes
the ends of the IS. Each type of element is named by giving it the
prefix IS followed by a number. In E. coli several copies of different IS elements have been observed; some of their properties.
Transposable elements also can contain genes other than
those required for transposition (for example, antibiotic resistance or toxin genes). These elements often are called composite
transposons or elements. Complete agreement about the nomenclature of transposable elements has not yet been reached. Sometimes transposable elements are called transposons when they
have extra genes, and insertion sequences when they lack these.
Composite transposons often consist of a central region containing the extra genes, flanked on both sides by IS elements that are identical or very similar in sequence.Many composite transposons are simpler in organization. They are bounded
by short inverted repeats, and the coding region contains both
transposition genes and the extra genes. It is believed that composite transposons are formed when two IS elements associate
with a central segment containing one or more genes. This association could arise if an IS element replicates and moves only a
gene or two down the chromosome. Composite transposon names
begin with the prefix Tn.
The process of transposition in procaryotes involves a series of
events, including self-replication and recombinational processes.
Typically in bacteria, the original transposon remains at the
parental site on the chromosome, while a replicated copy inserts at
the target DNA. This is called replicative transposition. Target sites are specific sequences about five to nine base pairs
long. When a transposon inserts at a target site, the target sequence
is duplicated so that short, direct-sequence repeats flank the transposon’s terminal inverted repeats. This can be seen where the five base pair target sequence moves to both
ends of the transposon and retains the same orientation.
The transposition of the Tn3 transposon is a well-studied example of replicative transposition. In the first stage the plasmid containing the Tn3 transposon fuses with the target plasmid to form a cointegrate molecule.
This process requires the Tn3 transposon enzyme coded for by the tnpA gene. Note
that the cointegrate has two copies of the Tn3transposon. In the
second stage the cointegrate is resolved to yield two plasmids, each
with a copy of the transposon. Resolution involves a crossover at the two ressites and is catalyzed by
a resolvase enzyme coded for by the tnp Rgene.
Transposable elements produce a variety of important effects.
They can insert within a gene to cause a mutation or stimulate
DNA rearrangement, leading to deletions of genetic material. If a
transposon insertion produces an obvious phenotypic change, the
gene can be tracked by following this altered phenotype. One can
fragment the genome and isolate the mutated fragment, thereby
partially purifying the gene. Thus transposons may be used to purify genes and study their functions. Because some transposons
carry stop codons or termination sequences, they may block translation or transcription. Other elements carry promoters and thus
activate genes near the point of insertion. Eucaryotic genes as well
as procaryotic genes can be turned on and off by transposon movement. Transposons also are located within plasmids and participate in such processes as plasmid fusion and the insertion of F
plasmids into the E. coli chromosome.
In the previous discussion of plasmids, it was noted that an R
plasmid can carry genes for resistance to several drugs. Transposons
have antibiotic resistance genes and play a major role in generating
these plasmids. Consequently the existence of these elements causes serious problems in the treatment of disease. Since plasmids can contain several different target sites, transposons will move between
them; thus plasmids act as both the source and the target for transposons with resistance genes. In fact, multiple drug resistance plasmids probably often arise from transposon accumulation on a single
plasmid. Because transposons also move between
plasmids and primary chromosomes, drug resistance genes can exchange between plasmids and chromosomes, resulting in the further
spread of antibiotic resistance.
Some transposons bear transfer genes and can move between bacteria through the process of conjugation, as discussed in the next section. A well-studied example of such a
conjugative transposon is Tn916 from Enterococcus faecalis.Although Tn916 cannot replicate autonomously, it will
transfer itself from E. faecalisto a variety of recipients and integrate into their chromosomes. Because it carries a gene for
tetracycline resistance, this conjugative transposon also
spreads drug resistance.
Please Refer This Link to:
en.wikipedia.org/wiki/Transposable_element
www.nature.com/scitable/topicpage/transposons-the-jumping-genes-518
www.nature.com/.../transposons-or-jumping-genes-not-junk-dna-1211
www.ndsu.edu/pubweb/~mcclean/plsc431/transelem/trans1.htm
www.bx.psu.edu/~ross/workmg/TranspositionCh9.htm
www.thefreedictionary.com/Transposable+element
www.ndsu.edu/pubweb/~mcclean/plsc431/transelem/trans5.htm
www.slideshare.net/ambicaparthasarathi/transposable-elements


Transposable Elements:
The chromosomes of bacteria, viruses, and eucaryotic cells contain pieces of DNA that move around the genome. Such movement
is called transposition. DNA segments that carry the genes required for this process and consequently move about chromosomes are transposable elements or transposons.Unlike other
processes that reorganize DNA, transposition does not require extensive areas of homology between the transposon and its destination site. Transposons behave somewhat like lysogenic prophages
except that they originate in one chromosomal
location and can move to a different location in the same chromosome. Transposable elements differ from phages in lacking a virus
life cycle and from plasmids in being unable to reproduce autonomously and to exist apart from the chromosome. They were
first discovered in the 1940s by Barbara McClintock during her
studies on maize genetics (a discovery that won her the Nobel
prize in 1983). They have been most intensely studied in bacteria.
The simplest transposable elements are insertion sequences
or IS elements . An IS element is a short sequence
of DNA (around 750 to 1,600 base pairs [bp] in length) containing only the genes for those enzymes required for its transposition and bounded at both ends by identical or very similar sequences of nucleotides in reversed orientation known as inverted
repeats . Inverted repeats are usually about 15 to 25
base pairs long and vary among IS elements so that each type of
IS has its own characteristic inverted repeats. Between the inverted repeats is a gene that codes for an enzyme called transposase(and sometimes a gene for another essential protein). This
enzyme is required for transposition and accurately recognizes
the ends of the IS. Each type of element is named by giving it the
prefix IS followed by a number. In E. coli several copies of different IS elements have been observed; some of their properties.
Transposable elements also can contain genes other than
those required for transposition (for example, antibiotic resistance or toxin genes). These elements often are called composite
transposons or elements. Complete agreement about the nomenclature of transposable elements has not yet been reached. Sometimes transposable elements are called transposons when they
have extra genes, and insertion sequences when they lack these.
Composite transposons often consist of a central region containing the extra genes, flanked on both sides by IS elements that are identical or very similar in sequence.Many composite transposons are simpler in organization. They are bounded
by short inverted repeats, and the coding region contains both
transposition genes and the extra genes. It is believed that composite transposons are formed when two IS elements associate
with a central segment containing one or more genes. This association could arise if an IS element replicates and moves only a
gene or two down the chromosome. Composite transposon names
begin with the prefix Tn.
The process of transposition in procaryotes involves a series of
events, including self-replication and recombinational processes.
Typically in bacteria, the original transposon remains at the
parental site on the chromosome, while a replicated copy inserts at
the target DNA. This is called replicative transposition. Target sites are specific sequences about five to nine base pairs
long. When a transposon inserts at a target site, the target sequence
is duplicated so that short, direct-sequence repeats flank the transposon’s terminal inverted repeats. This can be seen where the five base pair target sequence moves to both
ends of the transposon and retains the same orientation.
The transposition of the Tn3 transposon is a well-studied example of replicative transposition. In the first stage the plasmid containing the Tn3 transposon fuses with the target plasmid to form a cointegrate molecule.
This process requires the Tn3 transposon enzyme coded for by the tnpA gene. Note
that the cointegrate has two copies of the Tn3transposon. In the
second stage the cointegrate is resolved to yield two plasmids, each
with a copy of the transposon. Resolution involves a crossover at the two ressites and is catalyzed by
a resolvase enzyme coded for by the tnp Rgene.
Transposable elements produce a variety of important effects.
They can insert within a gene to cause a mutation or stimulate
DNA rearrangement, leading to deletions of genetic material. If a
transposon insertion produces an obvious phenotypic change, the
gene can be tracked by following this altered phenotype. One can
fragment the genome and isolate the mutated fragment, thereby
partially purifying the gene. Thus transposons may be used to purify genes and study their functions. Because some transposons
carry stop codons or termination sequences, they may block translation or transcription. Other elements carry promoters and thus
activate genes near the point of insertion. Eucaryotic genes as well
as procaryotic genes can be turned on and off by transposon movement. Transposons also are located within plasmids and participate in such processes as plasmid fusion and the insertion of F
plasmids into the E. coli chromosome.
In the previous discussion of plasmids, it was noted that an R
plasmid can carry genes for resistance to several drugs. Transposons
have antibiotic resistance genes and play a major role in generating
these plasmids. Consequently the existence of these elements causes serious problems in the treatment of disease. Since plasmids can contain several different target sites, transposons will move between
them; thus plasmids act as both the source and the target for transposons with resistance genes. In fact, multiple drug resistance plasmids probably often arise from transposon accumulation on a single
plasmid. Because transposons also move between
plasmids and primary chromosomes, drug resistance genes can exchange between plasmids and chromosomes, resulting in the further
spread of antibiotic resistance.
Some transposons bear transfer genes and can move between bacteria through the process of conjugation, as discussed in the next section. A well-studied example of such a
conjugative transposon is Tn916 from Enterococcus faecalis.Although Tn916 cannot replicate autonomously, it will
transfer itself from E. faecalisto a variety of recipients and integrate into their chromosomes. Because it carries a gene for
tetracycline resistance, this conjugative transposon also
spreads drug resistance.
Please Refer This Link to:
en.wikipedia.org/wiki/Transposable_element
www.nature.com/scitable/topicpage/transposons-the-jumping-genes-518
www.nature.com/.../transposons-or-jumping-genes-not-junk-dna-1211
www.ndsu.edu/pubweb/~mcclean/plsc431/transelem/trans1.htm
www.bx.psu.edu/~ross/workmg/TranspositionCh9.htm
www.thefreedictionary.com/Transposable+element
www.ndsu.edu/pubweb/~mcclean/plsc431/transelem/trans5.htm
www.slideshare.net/ambicaparthasarathi/transposable-elements

Comments