MICROBIAL METABOLISM CONTD...
FERMENTATION:
In the absence of aerobic or anaerobic respiration, NADH is not
oxidized by the electron transport chain because no external electron
acceptor is available. Yet NADH produced in the glycolytic
pathway during the oxidation of glyceraldehyde 3-phosphate to
1,3-bisphosphoglycerate must still be oxidized back
to NAD . If NAD is not regenerated, the oxidation of glyceraldehyde
3-phosphate will cease and glycolysis will stop. Many
microorganisms solve this problem by slowing or stopping pyruvate
dehydrogenase activity and using pyruvate or one of its derivatives
as an electron and hydrogen acceptor for the reoxidation
of NADH in a fermentation process . This may lead
to the production of more ATP. The process is so effective that
some chemoorganoheterotrophs do not carry out respiration even
when oxygen or another exogenous acceptor is available. There
are many kinds of fermentations, and they often are characteristic
 Fig: Advantages of Microbial Fermentation
Fig: Advantages of Microbial Fermentation


of particular microbial groups . A few more common
fermentations are introduced here, and several others are discussed
at later points. Two unifying themes should be kept in
mind when microbial fermentations are examined: (1) NADH is
oxidized to NAD , and (2) the electron acceptor is often either
pyruvate or a pyruvate derivative. In fermentation the substrate is
partially oxidized, ATP is formed by substrate-level phosphorylation
only, and oxygen is not needed.
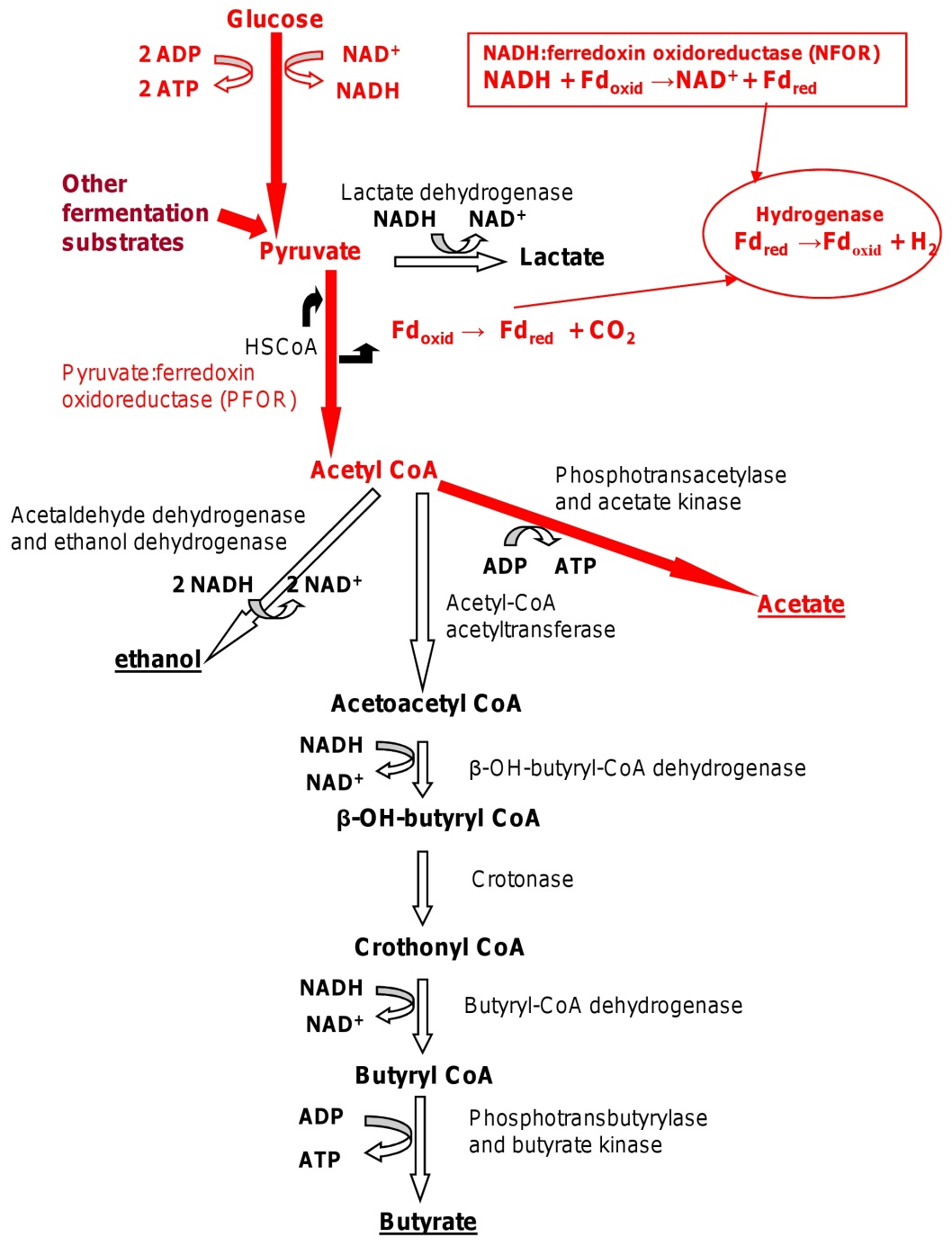
Many fungi and some bacteria, algae, and protozoa ferment
sugars to ethanol and CO2 in a process called alcoholic fermentation.
Pyruvate is decarboxylated to acetaldehyde, which is then
reduced to ethanol by alcohol dehydrogenase with NADH as the
electron donor . Lactic acid fermentation,
the reduction of pyruvate to lactate , is even
more common. It is present in bacteria (lactic acid bacteria, Bacillus),
algae (Chlorella), some water molds, protozoa, and even in animal skeletal muscle. Lactic acid fermenters can be separated
into two groups. Homolactic fermenters use the glycolytic
pathway and directly reduce almost all their pyruvate to lactate
with the enzyme lactate dehydrogenase. Heterolactic fermenters
form substantial amounts of products other than lactate;
many produce lactate, ethanol, and CO2 by way of the phosphoketolase pathway.
Alcoholic and lactic acid fermentations are quite useful. Alcoholic
fermentation by yeasts produces alcoholic beverages;
CO2 from this fermentation causes bread to rise. Lactic acid fermentation
can spoil foods, but also is used to make yogurt, sauerkraut,
and pickles. The role of fermentations in food production
is discussed in chapter 41.
Many bacteria, especially members of the family Enterobacteriaceae,
can metabolize pyruvate to formic acid and other products
in a process sometimes called the formic acid fermentation (figure
9.10, number 5). Formic acid may be converted to H2 and CO2 by
formic hydrogenlyase (a combination of at least two enzymes).
HCOOH→ CO2 H2
There are two types of formic acid fermentation. Mixed acid fermentation
results in the excretion of ethanol and a complex mixture
of acids, particularly acetic, lactic, succinic, and formic acids
(table 9.1). If formic hydrogenlyase is present, the formic acid
will be degraded to H2 and CO2. This pattern is seen in Escherichia,
Salmonella, Proteus, and other genera. The second
type, butanediol fermentation, is characteristic of Enterobacter,
Serratia, Erwinia, and some species of Bacillus (figure 9.10,
number 4). Pyruvate is converted to acetoin, which is then reduced
to 2,3-butanediol with NADH. A large amount of ethanol
is also produced, together with smaller amounts of the acids found in mixed acid fermentation.
Formic acid fermentations are very useful in identification of
members of the Enterobacteriaceae. Butanediol fermenters can
be distinguished from mixed acid fermenters in three ways:
1. The Voges-Proskauer test is a colorimetric procedure that detects the acetoin precursor of butanediol and is positive with butanediol fermenters but not with mixed
acid fermenters. The Voges-Proskauer test is used by both
the Enterotube II and API 20E microbial identification
systems to identify enteric bacteria.
2. Mixed acid fermenters produce four times more acidic
products than neutral ones, whereas butanediol
fermenters form mainly neutral products. Thus mixed
acid fermenters acidify incubation media to a much
greater extent. This is the basis of the methyl red test.
The test is positive only for mixed acid fermentation
because the pH drops below 4.4 and the color of the
indicator changes from yellow to red.
3 . CO2 and H2 arise in equal amounts from formic
hydrogenlyase activity during mixed acid fermentation.
Butanediol fermenters produce excess CO2 and the CO2/H2 ratio is closer to 5:1.
Formic acid fermenters sometimes generate ATP while reoxidizing
NADH. They use acetyl-CoA to synthesize acetyl phosphate,
which then donates its phosphate to ADP.
Acetyl-CoA Pi CoASH acetyl-P
Acetyl-P ADP acetate ATP
Microorganisms carry out fermentations other than those. Protozoa and fungi often ferment
sugars to lactate, ethanol, glycerol, succinate, formate, acetate,
 butanediol, and additional products.
butanediol, and additional products.
Substances other than sugars also are fermented by microorganisms.
For example, some members of the genus Clostridium prefer to ferment mixtures of
amino acids. Proteolytic clostridia such as the pathogens C.
sporogenes and C. botulinum will carry out the Stickland reaction
in which one amino acid is oxidized and a second amino
acid acts as the electron acceptor. Figure shows the way in
which alanine is oxidized and glycine reduced to produce acetate,
CO2, and NH3. Some ATP is formed from acetyl phosphate
by substrate-level phosphorylation, and the fermentation
is quite useful for growing in anaerobic, protein-rich environments.
The Stickland reaction is used to oxidize several amino
acids: alanine, leucine, isoleucine, valine, phenylalanine, tryptophan,
and histidine. Bacteria also ferment amino acids (e.g.,
alanine, glycine, glutamate, threonine, and arginine) by other
mechanisms. In addition to sugars and amino acids, organic
acids such as acetate, lactate, propionate, and citrate are fermented.
Some of these fermentations are of great practical importance.
For example, citrate can be converted to diacetyl and give flavor to fermented milk.

In the absence of aerobic or anaerobic respiration, NADH is not
oxidized by the electron transport chain because no external electron
acceptor is available. Yet NADH produced in the glycolytic
pathway during the oxidation of glyceraldehyde 3-phosphate to
1,3-bisphosphoglycerate must still be oxidized back
to NAD . If NAD is not regenerated, the oxidation of glyceraldehyde
3-phosphate will cease and glycolysis will stop. Many
microorganisms solve this problem by slowing or stopping pyruvate
dehydrogenase activity and using pyruvate or one of its derivatives
as an electron and hydrogen acceptor for the reoxidation
of NADH in a fermentation process . This may lead
to the production of more ATP. The process is so effective that
some chemoorganoheterotrophs do not carry out respiration even
when oxygen or another exogenous acceptor is available. There
are many kinds of fermentations, and they often are characteristic


fig:Lactic Acid Bacteria in Hydrogen-Producing Consortia: On Purpose or by Coincidence?

of particular microbial groups . A few more common
fermentations are introduced here, and several others are discussed
at later points. Two unifying themes should be kept in
mind when microbial fermentations are examined: (1) NADH is
oxidized to NAD , and (2) the electron acceptor is often either
pyruvate or a pyruvate derivative. In fermentation the substrate is
partially oxidized, ATP is formed by substrate-level phosphorylation
only, and oxygen is not needed.
Many fungi and some bacteria, algae, and protozoa ferment
sugars to ethanol and CO2 in a process called alcoholic fermentation.
Pyruvate is decarboxylated to acetaldehyde, which is then
reduced to ethanol by alcohol dehydrogenase with NADH as the
electron donor . Lactic acid fermentation,
the reduction of pyruvate to lactate , is even
more common. It is present in bacteria (lactic acid bacteria, Bacillus),
algae (Chlorella), some water molds, protozoa, and even in animal skeletal muscle. Lactic acid fermenters can be separated
into two groups. Homolactic fermenters use the glycolytic
pathway and directly reduce almost all their pyruvate to lactate
with the enzyme lactate dehydrogenase. Heterolactic fermenters
form substantial amounts of products other than lactate;
many produce lactate, ethanol, and CO2 by way of the phosphoketolase pathway.
Alcoholic and lactic acid fermentations are quite useful. Alcoholic
fermentation by yeasts produces alcoholic beverages;
CO2 from this fermentation causes bread to rise. Lactic acid fermentation
can spoil foods, but also is used to make yogurt, sauerkraut,
and pickles. The role of fermentations in food production
is discussed in chapter 41.
Many bacteria, especially members of the family Enterobacteriaceae,
can metabolize pyruvate to formic acid and other products
in a process sometimes called the formic acid fermentation (figure
9.10, number 5). Formic acid may be converted to H2 and CO2 by
formic hydrogenlyase (a combination of at least two enzymes).
HCOOH→ CO2 H2
There are two types of formic acid fermentation. Mixed acid fermentation
results in the excretion of ethanol and a complex mixture
of acids, particularly acetic, lactic, succinic, and formic acids
(table 9.1). If formic hydrogenlyase is present, the formic acid
will be degraded to H2 and CO2. This pattern is seen in Escherichia,
Salmonella, Proteus, and other genera. The second
type, butanediol fermentation, is characteristic of Enterobacter,
Serratia, Erwinia, and some species of Bacillus (figure 9.10,
number 4). Pyruvate is converted to acetoin, which is then reduced
to 2,3-butanediol with NADH. A large amount of ethanol
is also produced, together with smaller amounts of the acids found in mixed acid fermentation.
Formic acid fermentations are very useful in identification of
members of the Enterobacteriaceae. Butanediol fermenters can
be distinguished from mixed acid fermenters in three ways:
1. The Voges-Proskauer test is a colorimetric procedure that detects the acetoin precursor of butanediol and is positive with butanediol fermenters but not with mixed
acid fermenters. The Voges-Proskauer test is used by both
the Enterotube II and API 20E microbial identification
systems to identify enteric bacteria.
2. Mixed acid fermenters produce four times more acidic
products than neutral ones, whereas butanediol
fermenters form mainly neutral products. Thus mixed
acid fermenters acidify incubation media to a much
greater extent. This is the basis of the methyl red test.
The test is positive only for mixed acid fermentation
because the pH drops below 4.4 and the color of the
indicator changes from yellow to red.
3 . CO2 and H2 arise in equal amounts from formic
hydrogenlyase activity during mixed acid fermentation.
Butanediol fermenters produce excess CO2 and the CO2/H2 ratio is closer to 5:1.
Formic acid fermenters sometimes generate ATP while reoxidizing
NADH. They use acetyl-CoA to synthesize acetyl phosphate,
which then donates its phosphate to ADP.
Acetyl-CoA Pi CoASH acetyl-P
Acetyl-P ADP acetate ATP
Microorganisms carry out fermentations other than those. Protozoa and fungi often ferment
sugars to lactate, ethanol, glycerol, succinate, formate, acetate,
Substances other than sugars also are fermented by microorganisms.
For example, some members of the genus Clostridium prefer to ferment mixtures of
amino acids. Proteolytic clostridia such as the pathogens C.
sporogenes and C. botulinum will carry out the Stickland reaction
in which one amino acid is oxidized and a second amino
acid acts as the electron acceptor. Figure shows the way in
which alanine is oxidized and glycine reduced to produce acetate,
CO2, and NH3. Some ATP is formed from acetyl phosphate
by substrate-level phosphorylation, and the fermentation
is quite useful for growing in anaerobic, protein-rich environments.
The Stickland reaction is used to oxidize several amino
acids: alanine, leucine, isoleucine, valine, phenylalanine, tryptophan,
and histidine. Bacteria also ferment amino acids (e.g.,
alanine, glycine, glutamate, threonine, and arginine) by other
mechanisms. In addition to sugars and amino acids, organic
acids such as acetate, lactate, propionate, and citrate are fermented.
Some of these fermentations are of great practical importance.
For example, citrate can be converted to diacetyl and give flavor to fermented milk.

Comments